

クマは非常に順応性が高く万能な動物であり、森林、草原、山など広範囲に生息することができます。
さらに、彼らは雑食性で餌として消費される種の範囲は多岐に渡ります。それではなぜアフリカにはクマが存在しないのでしょうか?
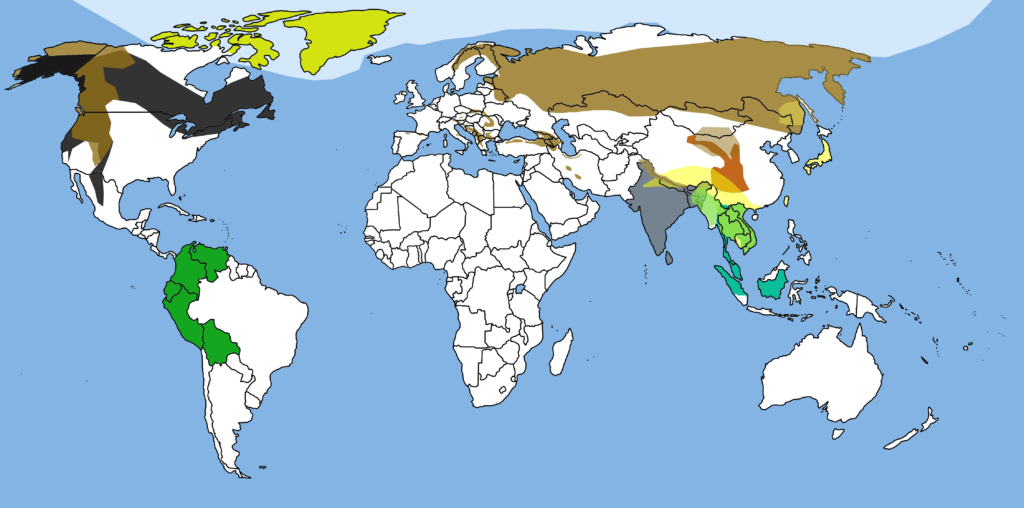
この記事の要約
- かつてアフリカには少なくとも2種類のクマが生息していた
- アトラスヒグマは19世紀まで北アフリカのアトラス山脈周辺に生息していたヒグマの亜種で、体長2.7m、体重450kg、植物食中心の雑食性だったが、人間による乱獲で1870年に絶滅した
- アグリオセリウム・アフリカヌムは中新世後期から鮮新世後期のサハラ以南アフリカに生息していたメガネグマ亜科で、体重最大750kg、植物質中心の雑食性で死骸あさりも行っていたが、サーベルタイガーなどとの生態的競争に敗れて約250万年前に絶滅した
- 現在アフリカにクマがいない理由はサハラ砂漠が地理的障壁となって新たなクマの南下を阻止し、アフリカの大型ネコ科動物との競争でクマ科が敗北したため
はじめに
一見するとクマがアフリカに存在しない論理的な理由はないように思えます。
クマは多様な環境に適応できる動物であり、多くの大陸に分布しているにもかかわらず、なぜアフリカでは見られないのでしょうか?
この疑問を解くために、まずは他の大陸の状況を比較してみましょう。
オーストラリアとアフリカの動物相の違い
オーストラリアにクマが存在しない理由は比較的単純です。 オーストラリアの哺乳動物相は独特であり、これは地質学的な歴史に起因しています。
オーストラリア大陸は何億年も前にゴンドワナ大陸と呼ばれる巨大大陸の一部でした。
この巨大大陸には現在のアフリカ、中東、南アメリカ、インド、南極、オーストラリア、ニュージーランドが含まれていたと考えられています。
中生代(2億5100万年前から1億4000万年前)にゴンドワナ大陸は分裂し、オーストラリア大陸は他から切り離されました。
それ以来、動植物はこの大陸で独自の進化を遂げ、その結果、有袋類を中心とした独特の生態系が形成されました。
一方、アフリカには他の大陸との関係を示すさまざまな現生哺乳類の分類群が存在しています。
アフリカに生息していた2種類のクマ
実は、かつてアフリカには2種類のクマが生息していました。
19世紀のアフリカ大陸北部、地中海沿いのアトラス山脈周辺に生息していた「アトラスヒグマ」(Ursus arctos crowtheri)と、中新世後期から鮮新世後期のサハラ以南のアフリカに生息していた「アグリオセリウム・アフリカヌム」(Agriotherium africanum)です。
これら2種のクマはどのようにアフリカに到達し、どのような生活を送り、そして絶滅したのでしょうか。この謎を解明するには、クマの進化の歴史を詳しく見ていく必要があります。
クマの起源と進化
最も初期の哺乳類は小さなげっ歯類ほどの大きさの生物でした。
クマの系統は肉食動物の起源と、裂肉歯と呼ばれる多くの肉食哺乳類において見られる肉や骨をハサミのように剪断する歯が発達する以前の暁新世の種「キモレステス」(Cimolesta)まで遡ることができます。

最初の真のクマはアライグマぐらいの大きさの哺乳類「パリクティス」(Parictis)から始まりました。
その後、3700万年前の中国の鉱床で発見された大型犬ほどの大きさの動物「セファロガレ」(Cephalogale)や「アンフィキノドン」(Amphicynodon)へと進化しました。
これらの種からさらに「フォベルシオン」(Phoberocyon)、「ピトキオン」(Pithocyon)、そして水生クマの「コロノモス」(Kolonomos)などに分岐していきました。
クマ科の3つの亜科
現在のクマ科は以下の3つの亜科に分けられます:
- ジャイアントパンダ亜科
- メガネグマ亜科
- クマ亜科(残りのすべてのクマが属します)
これら3つの系統の共通の祖先は2000万年前に生息していた「ウルサバス・エルメンシス」(Ursavus elmensis)です。
彼らはアジアで誕生し、やがて北米に広がっていきました。ウルサバスの体長は80cmほどで、おそらく現代のたぬきに似た姿をしており、雑食性で木登りが得意でした。

ジャイアントパンダ亜科
ジャイアントパンダはその祖先について長年議論の対象となっており、かつてはクマとして分類されていませんでした。
しかし今日では、ジャイアントパンダは700万年から800万年前に中国の雲南省に生息していた「アイルラルクトス」(Ailurarctos)の子孫であることが示されており、この種もウルサバスとの関連性を持っていました。
メガネグマ亜科
メガネグマ亜科は中新世中期に進化しました。
この時期、世界は乾燥化し、湿潤な森林は温帯林と低木地に取って代わられました。メガネグマ亜科の仲間はより特殊な肉食傾向を示すようになり、体も大きくなりました。
彼らは最初に北米に分散し、200万年前にパナマ地峡が形成されて南北両アメリカ大陸が陸続きになった時に南米へと進出しました。
主なメガネグマ亜科の種には、体重600kgの「ショートフェイスベア」(Arctodus simus)と、体重400kgで草食性の「フロリダホラアナグマ」(Tremarctos floridanus)がいます。
これら2つの種は目覚ましい成功を収め、わずか1万年前まで生き残っていました。
しかし、メガネグマ亜科は大型ネコ科動物との競争激化と更新世後期の大型草食動物の絶滅に直面しました。
さらに、彼らの絶滅は人類の出現とヒグマなどのクマ亜科の到来とも時期が一致しています。
現在、メガネグマ亜科として残っているのは南アメリカのメガネグマ(スペクタクルドベア)の1種だけです。
クマ亜科
クマ亜科は「オーヴェルニュグマ」(Ursus minimus)から500万年前に進化しましたが、150万年前の氷河期に大きな変化と様々な分岐を遂げました。
例えば、ユーラシア大陸のクマは大きさと俊敏さよりも、緩やかな動きと雑食性を進化の方向として選びました。
こうしてオーヴェルニュグマは現代のマレーグマ、ナマケグマ、アメリカグマ、ツキノワグマ、ヒグマ、ホッキョクグマなどの種を生み出しました。
オーヴェルニュグマはツキノワグマに最も似ていたと言われています。
120万から280万年前にオーヴェルニュグマからツキノワグマとヒグマが分岐し、これら2つのグループはヨーロッパとアジアに広がりました。
さらに氷期にシベリアとアラスカの間に形成されたベーリング陸橋を渡って北米に進出しました。
最も最近に進化した種はホッキョクグマで、約20万年前にヒグマの系統から分化しました。
極地という特殊な環境に生息するホッキョクグマはほぼ完全な肉食性を示しますが、その祖先であるヒグマは雑食性です。
このように、ヨーロッパ、アジア、アメリカ大陸でクマの進化の波が起こったことがわかります。
気候変動と地理的な連結部の形成により、クマは主要大陸へと分散することができました。
アメリカ大陸では、メガネグマ亜科の支配的地位は更新世にユーラシア大陸からのクマ亜科の侵入によって取って代わられました。
アトラスヒグマ:北アフリカの絶滅種
アフリカにもクマ亜科の一種であるヒグマが侵入していました。
アトラスヒグマは北アフリカに生息していたヒグマの亜種です。
この種は現代まで生き残ったアフリカ唯一の在来種のクマで、かつてはモロッコからリビアに至るアトラス山脈とその近隣地域に生息していました。
アトラスヒグマはわずかに明るい毛皮を持ち、中新世のクマを彷彿とさせる短い顔をしていました。
彼らの体長は2.7m、体重450kgに達したと考えられています。

アトラスヒグマの生態は他のヒグマと同様であると推定され、木の根やどんぐり、木の実などを食べていたようです。
主に植物食であったと言われていますが、今日のほとんどのクマは雑食動物であるため、アトラスヒグマも肉を食べることができ、おそらく小型の哺乳類も捕食していたと考えられています。
しかし、この種は現代の銃器が開発されてまもなく絶滅しました。
人間による乱獲がアトラスヒグマの減少の一因となった可能性があります。
最後の個体は1870年にモロッコ北部のテトゥアン山脈でハンターによって殺害されたと記録されています。
アグリオセリウム・アフリカヌム:サハラ以南の古代クマ
サハラ以南のアフリカでもクマの化石が発見されています。
これらの骨は中新世後期から鮮新世後期のメガネグマ亜科の1種に分類される「アグリオセリウム・アフリカヌム」のものです。
アグリオセリウム属の化石はアフリカだけでなく北米やユーラシアでも発見されています。
この属は非常に繁栄し、少なくとも約1160万年前から250万年前まで存続しました。
体重は最大750キログラムで、現生クマのほとんどよりも大きかったと考えられています。
アグリオセリウム属は他のクマ類よりも脚が長く顔が短く、体格も軽めでした。
幅広で短い顎は、強大な咬合力を生み出すことができました。

骨格に関する研究により、アグリオセリウム属は獲物を追いかけるような積極的な狩りに必要な四肢の強さやスピードを持っていなかったことが示されています。
また、歯や顎、歯の摩耗パターンの分析により、アグリオセリウム属は植物質を多く食べる雑食動物であることが判明しました。
炭素同位体の分析結果からは、現代のヒグマの一部の個体群と同様に、かなりの量の動物質も食べていたことがわかっています。
アグリオセリウム属は季節に応じて果物や無脊椎動物を食べるとともに、死骸をあさるために他の捕食者を追い払うという食性に特化していた可能性があります。
当時のアフリカやユーラシア、北アメリカには、サーベルタイガーやイヌ科のエピキオンなどの強力な肉食哺乳類が生息していました。
そのような優勢な捕食者たちが支配する環境で獲物を盗み、自らを守るためには、アグリオセリウム属の非常に大きな体格が必要だったと推測されます。
しかし、結果的にこの種は生態的競争に敗れ絶滅したと考えられています。
これがサハラ以南のアフリカにクマがいない理由を説明する重要な鍵となっています。
クマ科の拡散と障壁
クマの進化の多くはアジアや北アメリカで起こり、その結果生じた種は東西いずれかの方向に拡散しました。
アメリカ大陸ではパナマ地峡の形成後に南へと進出し、現在でもアンデス山脈沿いにメガネグマが生息しています。
アジアではジャイアントパンダが竹林で繁栄し、ツキノワグマが東南アジアを横断して南下し、ナマケグマがインドに進出しました。
そして、アトラスヒグマがサハラ砂漠の北のアトラス山脈沿いに生息していましたが、それより南には分布を広げることができませんでした。
サハラ砂漠は多くの種にとって南方への拡大に対する大きな障壁として機能しました。
この地理的障壁は過去百万年間、現生種のクマによるアフリカ南部への進出を阻んできました。先に述べたように、サハラ以南のアフリカでアグリオセリウム・アフリカヌムの化石が発見されていますが、この種は新たな分類群を生み出すことなく絶滅したようです。
これは生態的競争が絶滅の要因であったことを示唆しています。

競争に負けたクマ科
この状況をより深く理解するには、食肉目の進化を全体として考察する必要があります。
食肉目の祖先は暁新世から始新世中期(約6500万年前から4800万年前)に生息していた「ミアキス」と呼ばれる小型捕食動物です。
この動物はすぐに「イヌ型亜目」と「ネコ型亜目」の二つのグループに分化しました。
ネコ型亜目には全てのネコ科動物が含まれており、主にアフリカとアジアで進化的成功を収めました。
一方、イヌ、アライグマ、イタチ、アザラシ、そしてクマを含むイヌ型亜目は北米で最も繁栄し、そこからヨーロッパへと拡大していきました。
そのため、クマの系統は主に北半球で栄え、アフリカを通って南に効果的に拡散することができませんでした。
そして、アフリカに進出を試みたクマ科は、大型ネコ科動物のような優勢な捕食者との生態的競争に敗れたのです。
今日では、クマ科やイヌ科が多く生息する地域ではネコ科動物は比較的少なく、その逆も同様です。
まとめ
アフリカ大陸にクマが存在しない理由は、地理的障壁と生態的競争の組み合わせによるものだと考えられます。
北アフリカに生息していたアトラスヒグマは人間の狩猟により絶滅し、サハラ以南のアフリカに古代に生息していたアグリオセリウム・アフリカヌムは他の捕食者との競争に敗れました。
サハラ砂漠という強力な地理的障壁が、北からの新たなクマ種の侵入を妨げ、結果としてアフリカ大陸の大部分にクマが存在しない状況が生まれたのです。
ただし、これまでの説明は現在の生物地理学的知見と化石記録に基づくものであり、完全なものではありません。
近年の分類学と遺伝子検査技術の発達により、進化理論は飛躍的に進歩し続けており、将来的に新たな発見がこの説明に修正を加える可能性も十分にあります。
関連記事:クマ被害が増加している理由がわかるおすすめ本 | ジオチャン
参考:Why are there barely any bears in Africa? – Inside Ecology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
コメント